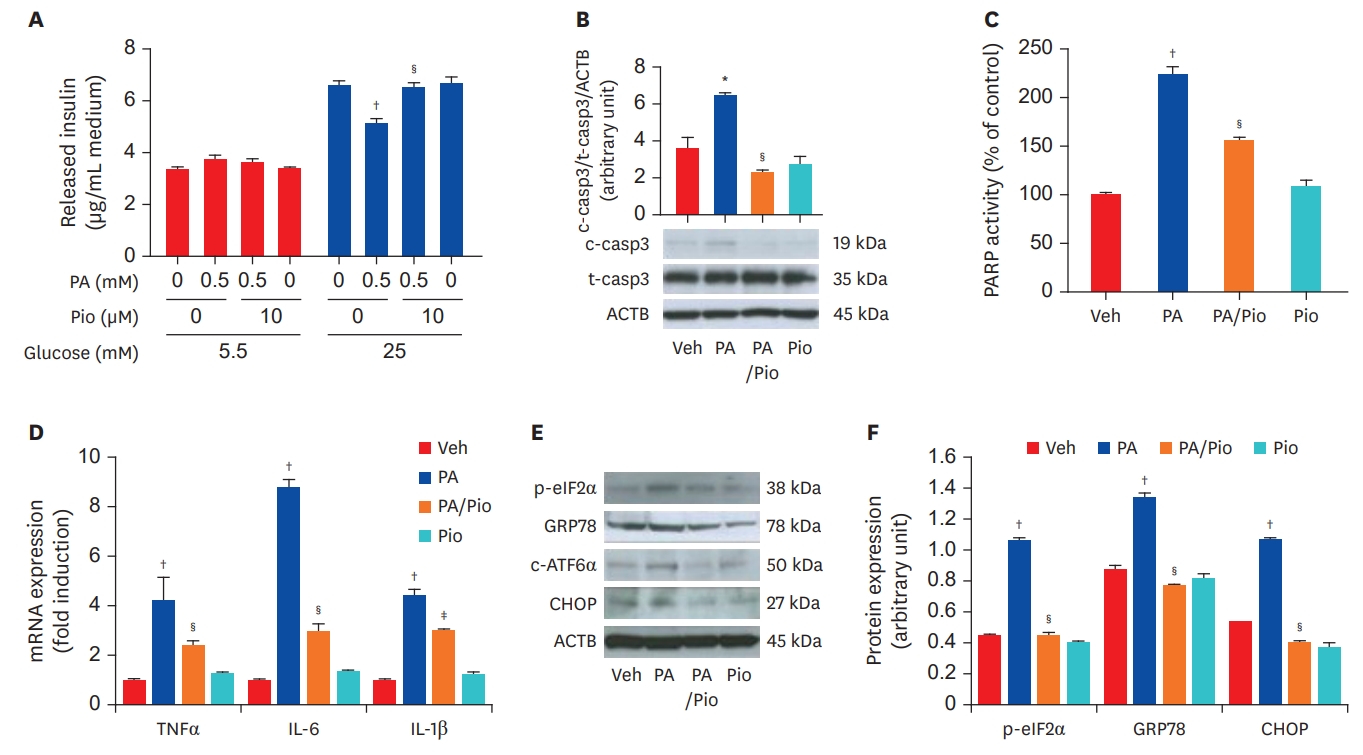
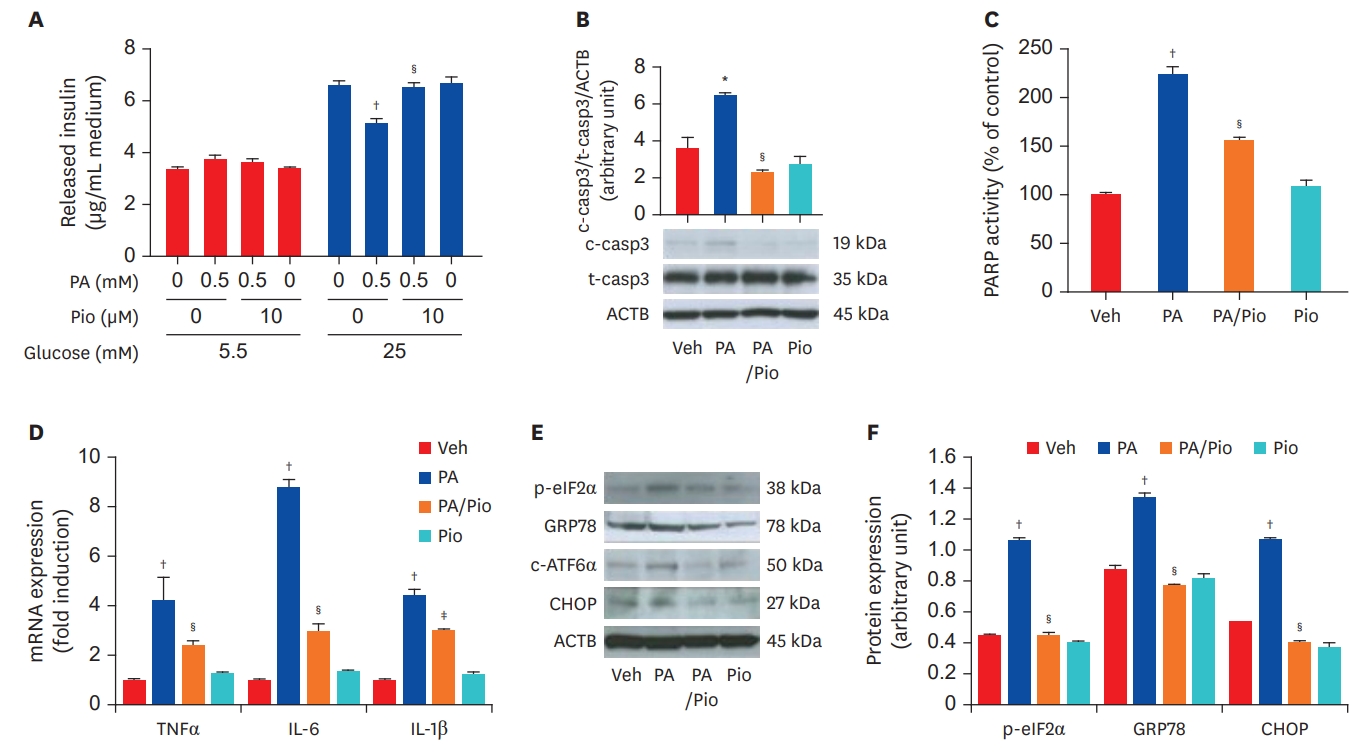
Division of Endocrinology and Metabolism, Department of Internal Medicine, Kangbuk Samsung Hospital, Sungkyunkwan University School of Medicine, Seoul, Korea
Copyright © 2021 Korean Society of Cardiovascular Disease Prevention; Korean Society of Cardiovascular Pharmacotherapy.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.

 PubReader
PubReader ePub Link
ePub Link Cite
Cite